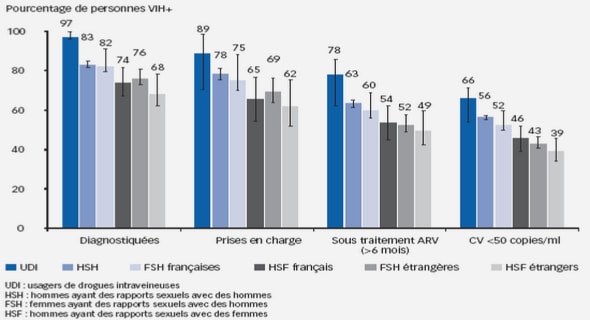
Le virus de l’hépatite C
Historique de la maladie
La découverte de l’hépatite B (VHB), en 1968 par Baruch Blumberg, et de celle de l’hépatite A (VHA), en 1973 par Stephen Feinstone, ont permis de mettre en évidence l’existence d’une « hépatite non-A non-B » chez des patients transfusés. En effet, en 1975, l’équipe de Feinstone a démontré, à partir d’une cohorte de vingt-deux patients ayant une hépatite associée à la transfusion, qu’au moins une partie de ces hépatites répondait négatif à des tests de sérologie spécifiques du VHA et du VHB. Les auteurs arrivaient à la conclusion, qu’au moins un autre agent infectieux était responsable d’hépatites transmissibles par voie parentérale. Ces hépatites prirent alors le nom « hépatite non-A non-B ». (3) En 1978, Alter et al. ont démontré que l’agent responsable de ces hépatites était transmissible en le transmettant à des chimpanzés. (4) C’est seulement en 1989, grâce à des techniques de biologie moléculaire, que l’équipe de Michael Houghton a réussi à isoler le clone d’un nouveau virus responsable d’hépatite non-A non-B qui a été baptisé « virus de l’hépatite C ». Grâce à cette découverte, un premier test sérologique de dépistage a pu être développé permettant de sécuriser les dons de sang et démontrant l’ampleur de la propagation de la maladie. (5) Les tests de détection des anticorps anti-VHC ont permis de mieux comprendre la maladie notamment son implication dans les carcinomes hépatocellulaires et dans d’autres maladies telles que des lymphomes, mais aussi de pouvoir évaluer les réponses virales aux traitements mis en place. En 2005, l’isolement de clones a permis le séquençage de l’ensemble du génome du VHC. De nouvelles approches thérapeutiques ont alors pu être développées grâce à cette découverte.
Caractéristiques virologiques
Classification du virus
Le virus de l’hépatite C appartient au genre Hepacivirus de la famille des Flaviviridae. Ce genre a été créé spécialement pour lui et ses variants. La famille des Flaviviridae est également composée du genre Flavivirus dont l’un des composants est le virus de la fièvre jaune, du genre Pestivirus dont l’un des représentants est le virus de la diarrhée des bovins et du genre Pegivirus. (7,8) Le VHC est très variable, actuellement 8 génotypes différents du VHC et de nombreux sous-types ont été confirmés.
Structure virale
Le VHC est un virus qui n’a pu être visualisé en microscopie électronique qu’à partir de 2016 grâce à une équipe de l’Inserm de Tours. En effet, avant cette découverte, toutes les données concernant le VHC avaient été obtenues par biologie moléculaire.
Le VHC est un virus enveloppé mesurant 40 à 100 nanomètres de diamètre. La particule virale contient une nucléocapside, qui apparait décentrée et qui contient l’ARN viral. Cette nucléocapside est entourée d’une monocouche de phospholipides. La particule virale contient également du cholestérol et des triglycérides. L’enveloppe extérieure, composée de phospholipides, est le lieu d’ancrage des glycoprotéines E1 et E2 qui sont organisées sous forme d’hétérodimères. Des apolipoprotéines dérivées de l’hôte, notamment ApoE, ApoB, et ApoC1, sont retrouvées en périphérie de la particule. La particule virale circule donc dans le sang sous forme de lipoviroparticule, avec une composition lipidique similaire aux VLDL, ce qui la rend difficilement observable au microscope.
La variabilité de taille entre les virions est due au nombre de couches de lipides qui les composent. Cette composition lipidique aurait un rôle dans la morphologie et l’infectiosité du virus, ainsi que dans la protection des glycoprotéines d’enveloppes vis-à-vis de la réponse immunitaire de l’hôte.
Structure du génome
Le génome est constitué d’une molécule d’ARN simple brin linéaire de polarité positive composée d’environ 9,6 kb. Il comprend un cadre de lecture ouvert unique entouré de deux régions non codantes qui ont un rôle dans la traduction et dans la réplication de l’ARN viral. La traduction du cadre de lecture ouvert va donner une polyprotéine unique d’environ 3000 acides aminés.
Fonctions des protéines virales
La polyprotéine obtenue par la traduction du cadre de lecture est le précurseur des protéines virales structurales (protéine de capside C et les glycoprotéines E1 et E2) et non-structurales (viroporine p7, non-structurales NS2, NS3, NS4A, NS4B, NS5A, NS5B).
Cycle viral
Le cycle de réplication du virus de l’hépatite C est un processus complexe faisant intervenir de nombreux médiateurs d’origine virale ou cellulaire. Le VHC n’est pas capable de synthétiser de l’ADN et ne peut donc pas s’intégrer au génome de l’hôte. La production virale est de 10¹² virions par jour. La réplication virale est rapide : en trois jours, l’ensemble de la charge virale serait renouvelé. De nombreuses recherches ont été effectuées, afin de le caractériser entièrement, mais pendant de nombreuses années, l’absence de système de culture cellulaire a été un frein à l’étude du cycle de réplication viral du VHC.
L’entrée du VHC dans la cellule est un processus complexe, qui impliquerait une cascade d’interactions entre de nombreux facteurs. La particule virale est transportée par la circulation sanguine jusqu’à la membrane basolatérale de l’hépatocyte où elle est initialement capturée par l’HSPG (héparane sulfate protéoglycane) et par le récepteur aux LDL (Low Density Lipoprotein) grâce à leurs interactions avec des apolipoprotéines, notamment ApoE, présentes sur la particule virale. (15,25) L’interaction de la glycoprotéine d’enveloppe E2 avec le récepteur SR-BI (receptor scavenger classe B type 1) permettrait un changement de conformation de E2, et initierait la liaison de CD81 avec E2. Cette interaction CD81-E2 provoquerait le déclenchement de voies de transduction du signal, entrainant la migration latérale de la particule virale vers une jonction serrée. La liaison CD81- E2 permettrait également un changement de conformation de l’hétérodimère E1E2. Il y aurait alors interaction entre le complexe E1E2 et deux protéines de jonctions serrées, la claudine-1 et l’occludine, qui seraient essentielles pour l’induction de l’endocytose.
Après l’attachement et le déplacement au niveau d’une jonction serrée, la particule virale va pouvoir rentrer dans la cellule par endocytose médiée par la clathrine. Une étape d’acidification endosomale est nécessaire pour entrainer des changements conformationnels dans les protéines de fusion permettant la fusion de l’enveloppe virale avec la membrane endosomale. Le génome viral va alors pouvoir être libéré dans le cytoplasme.
Dans le cytoplasme, l’ARN va être traduit en une polyprotéine unique qui sera ensuite clivée par des protéases virales et cellulaires pour la formation des protéines structurales (core, E1, E2) et des protéines non structurales (p7, NS2, NS3, NS4A, NS4B, NS5A et NS5B). Ces protéines sont nécessaires pour la réplication et pour la formation des particules virales. (7) La réplication du génome implique tout d’abord un réarrangement des membranes intracellulaires pour former un réseau membranaire. Ce réseau contient l’intégralité du complexe de réplication du virus qui est composé des protéines non structurales NS3, NS4A, NS4B, NS5A et NS5B et de facteurs cellulaires. Il apporte une protection lors de la réplication vis-à-vis des protéases et nucléases cellulaires. Les protéines NS4B et NS5A seraient impliquées dans la biogenèse et la maintenance de ces réseaux membranaires.
Dans le complexe de réplication, la protéine NS5B, qui est l’ARN polymérase dépendante de l’ARN viral, aidée par la protéine NS3 (hélicase et protéase), va permettre la synthèse d’un brin d’ARN, de polarité négative, complémentaire du génome viral. La protéine NS4A est un cofacteur de la protéine NS3 lui permettant de s’ancrer à la membrane. Ce brin de polarité négative va ensuite servir de matrice pour la formation excessive de nouveaux brins d’ARN de polarité positive qui vont ensuite être traduits et répliqués ou encapsidés pour la formation de nouvelles particules virales. De nombreuses autres protéines et microARN (miR 122) seraient aussi impliqués dans la réplication du VHC.
L’assemblage s’effectue au niveau d’un site spécifique situé à proximité des gouttelettes lipidiques cytosoliques où les protéines de capside se sont regroupées au préalable. L’interaction des protéines de capside avec les gouttelettes lipidiques serait essentielle pour le recrutement des protéines virales au niveau du site d’assemblage. Le génome viral va être transporté vers les protéines de capside notamment grâce à l’intervention de la protéine NS5A qui aurait un rôle essentiel dans ce transfert. (27,28) Les protéines de capside vont s’assembler pour former une nucléocapside entourant le génome viral. Les particules vont ensuite bourgeonner dans la lumière du réticulum endoplasmique et acquérir l’enveloppe phospholipidique et les glycoprotéines hétérodimériques E1E2. L’interaction ApoE avec E1-E2 est nécessaire pour une étape après l’enveloppement et avant la libération. (27) La dernière étape de la morphogenèse du VHC implique la voie de sécrétion des VLDL de l’hôte au cours de laquelle les virions du VHC s’associent aux VLDL ou à leurs composants pour former des lipoviroparticules avant leur libération. De nombreuses protéines impliquées dans la voie de biogenèse des VLDL semblent avoir un rôle dans le cycle de réplication virale, notamment ApoE qui a un rôle essentiel dans l’entrée et l’assemblage du virus. De nombreux autres facteurs hôtes interviendraient dans l’assemblage du VHC.
Le VHC peut également se transmettre entre cellules. Lors d’une infection chronique, le virus présent dans un hépatocyte infecté peut migrer vers un hépatocyte naïf. Ce passage de cellule à cellule permet la persistance de l’infection malgré la présence d’anticorps anti-VHC ou d’immunoglobulines. Une étude a démontré que cette méthode de dissémination serait responsable d’une résistance aux traitements. (25) Le VHC a été retrouvé dans des tissus autres que le foie entrainant des manifestations extra-hépatiques. Ces manifestations seront abordées par la suite.
Histoire naturelle de la maladie
Hépatite aiguë
La période d’incubation de l’hépatite C est de deux semaines à six mois. L’hépatite aiguë est dans la plupart des cas asymptomatique ou paucisymptomatique. Les symptômes pouvant être rencontrés sont de l’asthénie, de la fièvre, des nausées, une anorexie, des douleurs abdominales, des urines foncées ou encore un ictère. Les cas d’hépatite fulminante sont exceptionnels. La principale complication est le passage à la chronicité. En effet dans 70% des cas, l’hépatite aiguë va se chroniciser. Dans 30% des cas, le système immunitaire du malade réussit à se débarrasser du virus et permet la guérison. La guérison est affirmée par l’absence de détection du virus dans le sang au-delà de 6 mois après l’infection aiguë et la présence d’anticorps anti-VHC.
Hépatite chronique
Le virus de l’hépatite C n’a pas d’effet cytopathogène direct sur l’hépatocyte. C’est la réaction immunitaire de l’hôte qui entraine la cytolyse des hépatocytes, se caractérisant par l’apparition de lésions nécro-inflammatoires, d’un tissu cicatriciel fibreux qui se traduit, au fur et à mesure du temps, par une fibrose du foie. Le développement de la maladie est silencieux, et dans 20% des cas, la fibrose évolue en cirrhose en plusieurs décennies.
Les facteurs de risque favorisant la progression de la fibrose en cirrhose sont l’âge au moment de la contamination (patients âgés de plus de 40 ans), le sexe masculin, la présence de comorbidités (maladie hépatique préexistante, un syndrome métabolique : l’obésité, le diabète de type 2 même au stade de résistance à l’insuline), la co-infection par le VHB ou le VIH, la consommation d’alcool, de cannabis ou de tabac, le génotype viral (progression plus rapide avec le génotype de type 3 et moins rapide avec le génotype 1), des facteurs génétiques de l’hôte ou encore la prise de traitements immunosuppresseurs.
Le risque de décompensation de la cirrhose augmente de 3 à 6% par an, se caractérisant par une hypertension portale et/ou par une insuffisance hépatocellulaire. L’hypertension portale entraine des varices gastroœsophagiennes avec un risque d’hémorragie variqueuse, et de formation d’ascite. L’insuffisance hépatocellulaire peut être responsable de troubles de la coagulation, d’ictère et d’encéphalopathie hépatique. Dans l’année suivant la cirrhose décompensée, le risque de mortalité est compris entre 15 et 20%. (30) Le risque d’évolution de la cirrhose vers le carcinome hépatocellulaire (CHC) augmente de 3 à 8% par an. Le cancer du foie est le sixième cancer le plus fréquent dans le monde chez l’homme, et le quatrième en terme de mortalité par cancer. Environ un tiers des cancers de foie serait dû au VHC. L’apparition d’un CHC au stade de fibrose chez un patient atteint du VHC est très rare, c’est surtout quand le foie est au stade cirrhotique que le risque de développer un CHC est augmenté.
|
Table des matières
Introduction
1. Le virus de l’hépatite C
1.1 Historique de la maladie
1.2 Caractéristiques virologiques
1.2.1 Classification du virus
1.2.2 Structure virale
1.2.3 Structure du génome
1.2.4 Fonctions des protéines virales
1.2.5 Cycle viral
1.3 Histoire naturelle de la maladie
1.3.1 Hépatite aiguë
1.3.2 Hépatite chronique
1.3.3 Manifestations extra-hépatiques du VHC
1.4 Modes de transmission de l’hépatite C
1.4.1 Transmission horizontale
1.4.2 Transmission verticale
1.5 Epidémiologie
1.6 Dépistage et diagnostic de l’infection par le VHC
1.6.1 Stratégie de dépistage actuelle de l’infection par le VHC en France
1.6.2 Dépistage universel
1.6.3 Méthodes d’analyses
1.6.4 Test Rapide d’Orientation Diagnostique (TROD)
1.6.5 Campagnes de dépistage et de prévention
2. Prévention du risque de transmission du VHC
2.1 Politique de réduction des risques chez les toxicomanes
2.2 Politique de réduction des risques chez les populations migrantes
2.3 Sécurisation des dons de sang et des médicaments dérivés du sang
2.4 Prévention du risque de transmission lors des soins et lors des actes de type tatouage, piercing, maquillage permanent
3. Traitements et stratégie de prise en charge de l’hépatite C
3.1 Historique des traitements dans la lutte contre le VHC
3.2 Association interféron a et ribavirine
3.2.1 L’interféron a-pégylé
3.2.2 La Ribavirine
3.3 Antiviraux à action directe (AAD)
3.3.1 Généralités sur les antiviraux d’action directe
3.3.2 Les inhibiteurs de la protéase NS3-4A
3.3.3 Les inhibiteurs de la protéine NS5A
3.3.4 Les inhibiteurs de la protéine NS5B
3.3.5 Spécialités
3.4 Stratégies de prise en charge de l’hépatite C
3.4.1 Prise en charge dans un parcours simplifié
3.4.2 Prise en charge dans le cadre d’un parcours spécialisé
3.4.3 Surveillance du patient après guérison
4. Etude sur la prise en charge de l’hépatite C à l’officine en Seine-Maritime
4.1 Méthodologie
4.2 Résultats
5. Discussion : rôles du pharmacien d’officine dans la prise en charge de l’infection par le virus de l’hépatite C
5.1 Les atouts du pharmacien d’officine
5.2 Missions du pharmacien d’officine
5.3 Rôle du pharmacien d’officine dans la prévention primaire
5.4 Place du pharmacien d’officine dans le dépistage
5.4.1 Informer les patients
5.4.2 Dépistage à l’aide de TROD en officine
5.5 Rôle du pharmacien d’officine dans la dispensation du traitement
5.5.1 Analyse pharmaceutique
5.5.2 Conseils nécessaires au bon usage des médicaments
5.5.3 Adhésion thérapeutique
5.5.4 Freins à la dispensation des AAD à l’officine
5.6 Rôle du pharmacien d’officine dans la prévention tertiaire
5.7 Pluridisciplinarité
Conclusion